 SZ nr 27–48/2024
SZ nr 27–48/2024z 30 czerwca 2024 r.
Fenomenalne organoidy
Ewa Biernacka
Organoidy to samoorganizujące się wielokomórkowe struktury trójwymiarowe, które w warunkach in vitro odzwierciedlają budowę organów lub guzów nowotworowych in vivo. Żywe modele części lub całości narządów ludzkich w 3D, w skali od mikrometrów do milimetrów, wyhodowane z tzw. indukowanych pluripotentnych komórek macierzystych (ang. induced Pluripotent Stem Cells, iPSC) to nowe narzędzia badawcze w biologii i medycynie. Stanowią jedynie dostępny, niekontrowersyjny etycznie model wczesnego rozwoju organów człowieka o dużym potencjale do zastosowania klinicznego. Powstają w wielu laboratoriach na świecie, również w IMDiK PAN, gdzie badane są organoidy mózgu i nowotworowe. O twórcach i potencjale naukowym organoidów mówi prof. dr hab. n. med. Leonora Bużańska, kierownik Zakładu Bioinżynierii Komórek Macierzystych i dyrektor w Instytucie Medycyny Doświadczalnej i Klinicznej w Warszawie im. Mirosława Mossakowskiego Polskiej Akademii Nauk (IMDiK PAN).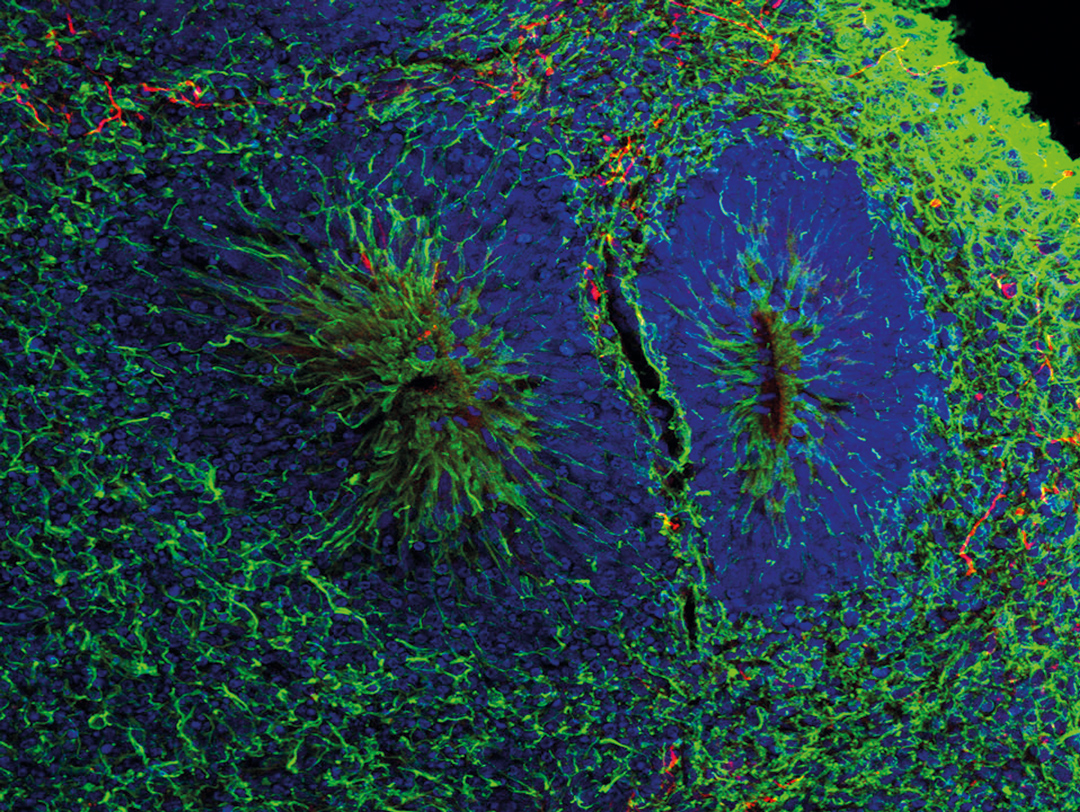
Fot. mgr inż. Michał Liput, IMDiK PAN
Pluripotentne komórki macierzyste
Najpierw odkryto niemal boską właściwość komórek macierzystych: zdolność do samoodnowy. Z dwóch komórek, które powstają po podziale, jedna, identyczna z komórką „matki”, pozostaje niezróżnicowana, druga różnicuje się w inne komórki organizmu. Zdolność do przekształcania się we wszystkie typy komórek organizmu (łącznie z linią komórek rozrodczych) mają pluripotentne komórki macierzyste. Podczas rozwoju człowieka występują w węźle zarodkowym blastocysty jako grupa komórek zwanych zarodkowymi komórkami macierzystymi (ang. Embryonic Stem Cells, ESC).
W jaki sposób zapłodnione jajo, pojedyncza komórka, jest zaprogramowana, by wytworzyć niezliczone wyspecjalizowane komórki, które łączą się w trójwymiarowe tkanki i narządy, badają od pokoleń wybitni embriolodzy, m.in. prof. Magdalena Żernicka-Goetz z University of Cambridge w Wlk. Brytanii oraz California University of Technology w USA, wychowanka i współpracowniczka profesorów: Andrzeja Tarkowskiego, laureata „Nagrody Japońskiej”, oraz noblistów: Martina Evansa i Johna Gurdona. Komórki ESC z zarodka ssaka (myszy) w 1981 r. wyizolował jako pierwszy i udowodnił, że można je hodować in vitro, Evans, zaś komórki zarodkowe ludzkie w 1998 r. James Thomson. Sir John Gurdon wiele lat wcześniej [1958], wykazał, że transfer jądra z komórki zróżnicowanej do cytoplazmy komórki jajowej (oocytu), powoduje odróżnicowanie (cofnięcie do wcześniejszego stadium rozwoju) przeniesionego materiału genetycznego do etapu, z którego można otrzymać cały organizm (klonowanie). Następnym krokiem milowym były badania Shinya Yamanaki, który zidentyfikował czynniki odpowiedzialne za odróżnicowanie i wprowadził je do fibroblastów (komórek zróżnicowanych z tkanki łącznej) myszy [2006], potem człowieka [2007]. Tak, bez niszczenia zarodka, uzyskał indukowane pluripotencjalne komórki macierzyste iPSC. Zastosowane czynniki Yamanaki (Oct4, Sox-2, Klf-4, c-Myc) „odmłodziły” komórkę bez zmiany zapisu genetycznego poprzez tzw. regulację epigenetyczną, powodującą zmianę aktywności określonych genów. Geny odpowiedzialne za pluripotentność zostały aktywowane, a typowe dla komórki zróżnicowanej wyciszone. John Gurdon oraz Shinya Yamanaka otrzymali Nagrodę Nobla [2012] za „udowodnienie, że komórki budujące dorosły organizm mogą zostać reprogramowane tak, że stają się pluripotentne”. iPSC, otrzymane na drodze reprogramowania, analogicznie do komórek macierzystych pobranych z rozwijającego się zarodka, mogą być różnicowane in vitro we wszystkie inne typy komórek, np. w neurony lub erytrocyty. Tak otrzymane, nie są kontrowersyjne etycznie, a metoda Yamanaki jest uniwersalna, do powtórzenia w każdym laboratorium.Narodziny organoidy jako struktury samoorganizującej się
Jak w klasycznym teatrze, w powstawaniu organoidy wszystko ma swoje miejsce i czas. Zapisane w kodzie genetycznym każdej komórki czynniki transkrypcyjne, na określonym etapie rozwoju są aktywowane lub wyciszane, a ich dostępność wpływa na transkrypcję genów odpowiedzialnych za zaprogramowany rozwój. Różnicowanie komórek pluripotentnych w określonym kierunku można nazwać procesem programowania, który w warunkach in vitro jest częściowo sterowany przez badacza, a częściowo determinowany przez wewnętrzny program rozwoju komórki.
Organoidy powstają dzięki unikatowej zdolności komórek iPSC do samoorganizacji. Jest to możliwe dzięki ekspresji białek adhezyjnych występujących w błonie komórkowej (kadheryn), które poprzez selektywną adhezję sterują początkowym procesem tworzenia konglomeratów komórkowych. Takie konglomeraty komórkowe (zwane koloniami, jeśli powstały z jednej komórki), wyrastają na żelu w warunkach przylegania do podłoża. Po przeniesieniu na powierzchnię nieadhezyjną lub do rotacyjnych bioreaktów, w zależności od warunków rozwoju (pożywka hodowlana z czynnikami wzrostu i różnicowania, podawanymi w odpowiednim czasie), samoorganizują się w populacje założycielskie tkanek i narządów, naśladując ich architekturę i funkcje. Organizację struktury przestrzennej o ustalonych osiach rozwoju wymusza różna orientacja osi podziałów komórkowych i gradienty czynników (morfogenów).
Rozkwit modeli
Pierwsze struktury o charakterze organoidów powstały w laboratoriach Sasai (kora mózgu w 2008 r.) i Hansa Cleversa (krypty jelitowe w 2009 r.). Zespół Sasai w zastosowanym protokole wykorzystał małe cząsteczki i cytokiny do specyficznego kierowania agregatów PSC w stronę neuronabłonka (struktury mózgowe). Zespół Cleversa wykazał, że pojedyncze komórki macierzyste, wyizolowane z krypt jelitowych, mają potencjał ponownego wzrostu całego nabłonka jelitowego. Organoidy oparte na PSC są obecnie modelami wątroby, serca, nerek, płuc, trzustki, prostaty i innych organów.
Pierwsze organoidy nowotworowe otrzymano w 2014 (rak prostaty) i 2015 r. (rak trzustki).
Organoidy mózgu
Organoidy mózgu odzwierciedlają w dużej części zarówno przestrzenną budowę warstwową, jak i procesy funkcjonalne najbardziej złożonej tkanki ludzkiej, która buduje mózg. Obecnie stosowane metody umożliwiają utrzymanie takich struktur w hodowli in vitro do czterech lat. Różnorodność komórkowa pojawia się w nich ok. 30. dnia od jej założenia, a architektura struktur kory mózgu po ok. 2. miesiącach. Dotyczy to organoidów mózgu z reprezentacją komórek i struktury typowej dla całego mózgu (ang. cerebral organoids), które powstają bez dodatkowych czynników kierunkujących rozwój. Dodanie czynników (small moleculs) wspierających regionalizację mózgu pozwala otrzymać organoidy typowe dla określonego jego obszaru, np. kory mózgu, prążkowia czy wzgórza. Takie struktury można łączyć ze sobą w tzw. assembloidy do badania oddziaływania między obszarami mózgu. Strategia otrzymywania organoidów typowych dla różnych części mózgu inspirowana jest wiedzą, jaką mamy na temat regionalizacji mózgu podczas wczesnego rozwoju oraz tworzenia gradientów czynników określonych jako morfogeny, które wyznaczają osie brzuszno-grzbietową i przednio-tylną.
Wiemy również, że organoidy ludzkie wszczepiane do kory mózgu gryzoni mogą być unaczynione, a neurony tworzą funkcjonalne połączenia z siecią neuronalną otaczającej tkanki.
Yoshike Sasai, Instytut Riken w Kobe, 2008
Jako pierwszy pokazał wbudowaną w komórki prekursorowe możliwość tworzenia złożonych struktur kory mózgu. Wykorzystując kreatywnie warunki hodowli i wgląd w działanie sygnałów biologicznych rządzących rozwojem tkanek, naśladował tajemnicze procesy rozwoju mózgu w laboratorium. Był mistrzem w rozszyfrowywaniu kodu, dzięki któremu komórki uczą się o swoim miejscu w rozwijającym się zarodku. Przetestował ten kod na kulturach ludzkich ESC i poinstruował te niewyspecjalizowane komórki, aby stały się określonymi typami neuronów. Otrzymał komórki nerwowe znajdujące się w różnych częściach przodomózgowia, zmieniając stężenie morfogenów, cząsteczek kierujących tworzeniem wzorów tkanek. Ostrożnie dodając czynniki wzrostu, w zdefiniowanej kolejności i określonym czasie rozwoju, nakłonił organoidy do uformowania przedniej lub tylnej części kory mózgu – wprowadził metody instrukcji komórek ES, aby stały się neuronami korowymi. Powstało wspaniałe narzędzie do badania prawidłowego i nieprawidłowego rozwoju ludzkiego mózgu. Grupa Sasai była pionierem trójwymiarowej hodowli in vitro struktur kielicha (pęcherzyka) ocznego i kory rozwijającego się mózgu człowieka.
Madeline Lancaster
Biolożka rozwojowa w Laboratorium Biologii Molekularnej Medical Research Council w Cambridge w Wielkiej Brytanii w 2013 r. w grupie prof. Jürgena Knoblicha w Wiedniu otrzymała słynną organoidę z zalążkami całego mózgu: Cerebral Organoid, co zapoczątkowało badania nad modelowaniem rozwoju ludzkiego mózgu i schorzeń neurologicznych in vitro.
Genomika, transkryptomika i proteomika w badaniu na organoidach
Transkryptomika z użyciem narzędzi molekularnych, jak single-cell transcriptomics (sCT), identyfikuje ekspresję genów w komórce, wskazuje które są aktywne (uległy transkrypcji) i nieaktywne. Forma przestrzenna sCT pokazuję, gdzie w organoidzie znajdują się te komórki, a genomika funkcjonalna umożliwia śledzenie ich losów (ang. lineage tracing).
Ekspresję białek wykrywają badania proteomiczne. Białka znakowane metodą immunocytochemii identyfikują strefy proliferacji neuralnych komórek macierzystych, pozwalają odróżniać neurony, astrocyty i inne komórki. Ekspresja białka Ki 67 identyfikuje komórki w trakcie podziału.
Doskonalenie organoidów
By dostarczyć organoidom więcej tlenu i składników, próbuje się je unaczyniać, indukując ekspresję białek i rozwój komórek odpowiedzialnych za powstawanie naczyń krwionośnych. Inny sposób to łączenie organoidy, np. przodomózgowia, z organoidą naczyń krwionośnych w assembloidzie. W laboratorium Freda Gage w Instytucie Salka w Kalifornii [Nature Biotechnology, 2018] po raz pierwszy wszczepiono organoidę do mózgu myszy, uzyskując infiltrację do niej naczyń krwionośnych. Wzbogacanie organoidów w komórki mikrogleju umożliwia badania schorzeń neurozapalnych i neuroprotekcyjnych.
Ogromny potencjał
Organoidy mają udział w badaniach biologii rozwoju człowieka. Umożliwiają poszukiwanie mechanizmów ewolucyjnych, które doprowadziły do różnic rozwojowych, które czynią nas ludźmi, podsumowanie kamieni milowych w rozwoju człowieka i manipulowanie procesami rozwojowymi w celu zrozumienia, w jaki sposób wyłonił się złożony ludzki mózg i co jest przyczyną chorób rozwojowych. Naśladują przestrzennie i czasowo złożone procesy organogenezy w warunkach laboratoryjnych, dają wgląd w powstawanie, rozwój i funkcje tkanek i organów ludzkiego organizmu, w mechanizmy komórkowe. Odtwarzają złożone sieci funkcjonalne i fenotypy chorób, procesy inicjacji zaburzeń, jak nadekspresję lub zbytnie hamowanie genów, brak równowagi w szlakach sygnałowych. Umożliwiają tworzenie modeli chorób neurorozwojowych (padaczka, m.in. zespół Draveta), neurodegeneracyjnych (Alzheimer, Parkinson) i neuropsychiatrycznych mózgu (autyzm, demencja, schizofrenia). Za pomocą edycji genomu zmienia się sekwencje w obrębie istniejących genów lub usuwa mutacje genetyczne. Tak powstają tzw. linie izogeniczne iPSC, stosowane do testowania nowych terapii. Planuje się otrzymywać populacje komórek kompetentnych terapeutycznie jako źródło komórek i tkanek do przeszczepów i terapii komórkowej w medycynie regeneracyjnej. Co więcej, zastosowanie organoidów do modelowania chorób in vitro ogranicza eksperymenty na zwierzętach.